1. Introduction
Antibody-drug conjugates (ADCs) are considered to be among the most promising approaches to treat cell specific disorders. By making use of a targeting moiety which binds to a molecular target, a (cytotoxic) drug can be delivered specifically to diseased cells. ADCs consist of a small molecule drug which is attached via a linker molecule to the targeting moiety, for which antibodies are mostly used. Unless an extracellularly cleavable linker is employed, the ADC should internalize into the target cell, traffic to the lysosomes where the low pH and presence of lysosomal enzymes such as Cathepsin B release the drug from the targeting moiety [
1,
2,
3,
4]. Only then the drug can reach its action site, such as the microtubules. Therefore, internalization speed and intracellular trafficking of an ADC may have considerable impact on the functionality and effectivity of an ADC [
5].
While conventional antibodies are often used as a targeting moiety, they come with several limitations: they are large in size and cannot easily extravasate and penetrate tissue to reach all the target cells [
6,
7]. Moreover, the internalization rate and intracellular fate of non-engineered antibodies may not be optimal to serve as targeting moiety for intracellular drug delivery [
8].
Recent studies have suggested that small sized (15 kDa) single domain antibodies (sdAbs) are promising candidate targeting moieties for intracellular drug delivery. The sdAbs used here are VHHs, the variable domain of heavy chain only antibodies derived from the camelid family [
9]. They can access and bind very small epitopes with high affinity and are able to reach small immunogenic pockets [
10]. An epidermal growth factor receptor (EGFR) targeted by a half-life extended biparatopic sdAb conjugated to a platinum prodrug efficiently inhibited the growth of EGFR positive tumors [
11]. In another recent study, a single dose administration of a half-life extended sdAb targeting the human epidermal growth factor receptor 2 (HER2) conjugated to Auristatin F demonstrated excellent efficacy in vivo and led to durable tumor remissions (>124 days) [
12]. Another study compared a monoclonal antibody and sdAbs targeting tumors expressing prostate-specific membrane antigen (PSMA) conjugated to a potent DNA-alkylating agent. The authors showed that when the internalization speed was similar, the in vivo efficacy of the smaller sdAb was exceeding the monoclonal antibody [
13]. These results are in line with the rationale that smaller sized targeting moieties such as sdAbs enable better drug delivery than full-sized antibodies.
Here, we selected sdAbs against the rat platelet derived growth factor β receptor (PDGFRβ) transmembrane receptor as a model target because it is reported to be overexpressed in different types of diseases, such as non-small-cell lung cancer, esophageal cancer, gastric cancer and human muscular dystrophy [
14,
15,
16,
17,
18]. For this proof of principle study, we aimed to select internalizing sdAbs with either high or low internalization rates and to combine them into bivalent and biparatopic formats and investigate their internalization kinetics and drug delivery capacity.
Interestingly, our results show that monovalent sdAbs, targeting the same receptor, can be selected with different internalization rates. After conjugation of the different sdAb formats to a cytotoxic drug, we demonstrated that the targeting moieties deliver the drugs to the lysosomal cell compartment and that targeting moieties with a higher internalization rate show improved in vitro efficacy compared to the targeting moieties with a lower internalization rate.
2. Materials and Methods
2.1. Cell Lines
The mouse squamous cell carcinoma cell line (SCCVII) was kindly provided by prof. dr. Lukas Mach, of the University of Natural Resources and Life Sciences in Vienna, Austria. These SCC cells were stably transfected with rat PDGFRβ-GFP (SCCVII-rPDGFRβ) and clone sorted for high GFP signal by FACS. The cell lines were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) (Lonza, Basel, Switzerland) with 100 units/mL streptomycin, 0.1 mg/mL penicillin and 10% fetal bovine serum (FBS). To maintain transgene expression, the cells were kept under selection pressure with 400 µg/mL G418 Sulfate (Thermo Fisher Scientific, Bleiswijk, The Netherlands). Cells were kept at 37 °C in a humidified atmosphere containing 5% CO2 and were repetitively tested negative for Mycoplasma.
2.2. Construction of Immune sdAb Phagemid Libraries
Immunization of two llamas was performed at Eurogentec (Seraing, Belgium). Animals received four injections of rat PDGFRβ protein ectodomain (25 μg per injection; Sino-Biological, Eschborn, Germany) with intervals of 2 weeks. Four weeks after the last interval, a final boost injection with SCCVII-rPDGFRβ (10
8 cells) was administered. Llama serum was obtained, and the total mRNA isolated from peripheral blood lymphocytes was transcribed to cDNA. Specific primers were used to amplify the variable domain of the heavy chain only antibodies, which were eventually ligated into a phagemid vector as described previously [
19]. Transformation of electrocompetent TG1 cells resulted in the generation of two libraries of approximately 10
6–10
7 clones, which were further used for phage display.
2.3. Evaluation of Anti-Rat PDGFRβ Immune Response in Llama Sera
Recombinant rat PDGFRβ (100 ng per well, Sino-Biological, Eschborn, Germany) was coated overnight at 4 °C in PBS in 96-well flat bottom plates (Nunc MaxiSorp™, Thermo Fisher Scientific, Bleiswijk, the Netherlands). Next day, plates were washed with PBS and blocked with blocking buffer (PBS, supplemented with 4% (w/v) skimmed milk powder (Merck Millipore, Darmstadt, Germany) for 1 h at room temperature (RT). Serial dilutions of pre-immune and immune sera were added in 2% MPBS (PBS, supplemented with 2% (w/v) skimmed milk powder (Merck Millipore, Darmstadt, Germany)). Detection of bound antibody was performed by incubation with rabbit anti-llama Ig antiserum (in 2% MPBS; Jackson Immunoresearch, West Grove, PA, USA) and horseradish peroxidase-conjugated donkey anti-rabbit Ig (in 2% MPBS; Jackson Immunoresearch, West Grove, PA, USA). All incubations were carried out for 1 h at RT and after every incubation, plates were washed four times with PBS. For quantification, 3,3′,5,5′-tetramethylbenzidine (TMB) substrate was added and the OD of the solution was measured at 450 nm.
2.4. Phage Display Selection of Internalizing Rat PDGFRβ Specific sdAbs
To isolate rat PDGFRβ sdAbs, two consecutive phage panning rounds were performed. Phages were first panned on purified protein, followed by a second panning round on SCCVII-rPDGFRβ cells. Phage selection in the first panning round was performed as described previously [
19] with the only difference being the amount of coated protein which was 0.05 µg/mL of PDGFRβ protein. In the second panning round, 2.5 × 10
5 SCCVII-rPDGFRβ cells were used. To skew selection towards sdAbs with internalizing ability, bio-panning was performed for 1 h at 37 °C, and the phages that bound to the membrane of SCCVII-rPDGFRβ cells were removed using an acid wash. Subsequently, the SCCVII-rPDGFRβ cells were lysed and the remaining internalized phages were used for
E. coli TG1 cells infection. A colony PCR was performed to determine the number of clones containing a sdAb insert. Identifying unique clones in the selected library was accomplished with HinfI digestion (Thermo Fisher Scientific, Bleiswijk, The Netherlands).
2.5. sdAbs Production and Purification
2.5.1. For Direct Use of the Periplasmic Fraction
In a deep well plate, 1.5 mL LB medium 0.2% (w/v) glucose with 100 μg/mL ampicillin was inoculated with E. coli TG1 containing the sdAb in the phagemid vector and incubated for 3 h at 37 °C at 180 rpm. The culture was then induced with 1 mM IPTG for overnighy sdAb production. Next morning, the culture was harvested by spinning down at 5000× g at 4 °C for 20 min. The pellet was resuspended in 100 µL PBS and freeze-thawed twice to release periplasmic content. The bacteria were then spun down at 5000× g at 4 °C for 20 min to collect the bacterial periplasm, which was then used directly in a binding assay on immobilized ectodomain.
2.5.2. For Purification
Initial productions and binding assays with the monomeric sdAbs were performed with the sdAbs in the original phagemid vector. The selected sdAb sequences were re-cloned in a modified pET21 to introduce a N-terminal pelB signal sequence and a C-terminal cysteine and EPEA tag. For productions, pre-cultures were made by inoculating E. coli BL21-DE3 Codonplus (Stragene, Bellingham, WA, USA) containing the sdAb in pET21 vector into 90 mL LB medium, 2% (w/v) glucose and 100 μg/mL ampicillin, which was incubated overnight at 37 °C at 180 rpm. The 5 L Eppendorf BioFlo®/CelliGen® 115 fermentor containing probes for dissolved oxygen (DO), pH and temperature measurements was prepared according to the manufacturer’s instructions and all measurements were monitored via the BioCommand program. A 1:100 dilution of the overnight pre-culture was added to 5 L Terrific Broth (TB) medium, supplemented with 17 mM KH2PO4, 72 mM K2HPO4, 0.1% (w/v) glucose, 100 μg/mL ampicillin and 1:10,000 anti-foam. Bacteria were grown at 37 °C and OD600 was monitored using a spectrophotometer until the desired OD600 of ±1 was reached. Temperature was then lowered to 25 °C and the bacteria were induced with 1 mM IPTG for overnight sdAb production. Next morning, the culture was harvested by spinning down the culture at 5000× g at 4 °C for 20 min. The pellet was resuspended in 750 mL PBS and freeze-thawed twice to release periplasmic content. The bacteria were then spun down at 5000× g at 4 °C for 30 min to collect the bacterial periplasm, which was then filtrated through a 0.45 µm filter.
Periplasmic fraction containing EPEA-tagged sdAb was purified on the ÄKTAXpress chromatography system using a 1 mL CaptureSelect™ C-tag column (Thermo Fisher Scientific, Bleiswijk, The Netherlands) and 2 × 5 mL HiTrap Desalting Columns (GE Healthcare, Hoevelaken, The Netherlands). Sample was loaded (1 mL/min) onto the C-tag column, after which bound sdAb was eluted using 2 mM Tris, 2 M MgCl2, pH 7 and buffer exchanged to PBS using the HiTrap Desalting columns. Purified fractions were loaded on with SDS-PAGE gels for purity assessment and stored at −80 °C.
2.6. Conjugations
2.6.1. sdAb—Maleimide—Fluorophore Conjugation
The free C-terminal cysteine in the sdAbs was used for site-directed maleimide-fluorophore conjugation using a procedure described elsewhere [
20]. Briefly, sdAbs were incubated with 2 molar equivalents of TCEP in borate buffer (25 mM sodium borate pH 8, 25 mM NaCl, 1 mM DTPA) at 37 °C for 2 h. The maleimide-IRDye800CW (LI-COR Biosciences, Lincoln, NE, USA) or maleimide-AlexaFluor647 (Thermo Fisher Scientific, Bleiswijk, The Netherlands) was added at 5–10 molar equivalents and incubated on ice for 1 h. The conjugates were purified from free fluorophore using two consecutive 2 mL Zeba spin desalting columns (Thermo Fisher Scientific, Bleiswijk, The Netherlands) which were pre-equilibrated with PBS. The amount of free dye in the samples was determined by SDS-PAGE. Upon gel electrophoresis, fluorescence was detected with an Odyssey infrared scanner at 700 nm or 800 nm. The degree of conjugation (DoC) was determined following the manufacturer’s protocol by measuring the absorbance at 280 and 650 nm for AlexaFluor647 and 280 and 800 nm for IRDye800 using a Nanodrop spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA).
2.6.2. sdAb—Lx—Auristatin F Conjugation
The free C-terminal cysteine in the sdAbs was used for site-directed Auristatin F conjugation using the Lx linker technology as described elsewhere [
21]. Briefly, sdAbs were incubated with 2 molar equivalents of TCEP in borate buffer (25 mM sodium borate pH 8, 25 mM NaCl, 1 mM DTPA) at 37 °C for 2 h. The AF-Lx-thiourea (prepared by mixing AF-Lx-I and 20 mM thiourea (1:1) at 37 °C for 2 h) was added to the reduced sdAbs and incubated at 37 °C for 1 h. The conjugates were purified using 10 kDa Amicon Ultra centrifugal filters (Merck Millipore, Darmstadt, Germany). The purity and degree of conjugation (DoC) of the conjugates was determined by SEC-MS.
2.7. Binding Assays
2.7.1. On Immobilized Ectodomain
Purified rat PDGFRβ was purchased from Sino-Biological (Eschborn, Germany) and coated overnight at 4 °C at 1 μg/mL in PBS in ELISA plates (Nunc MaxiSorp™, Thermo Fisher Scientific, Bleiswijk, the Netherlands). Next day, plates were washed with PBS and blocked with blocking buffer (PBS, supplemented with 4% (w/v) skimmed milk powder (Merck Millipore, Darmstadt, Germany)) for 1 h at RT. Next, three-fold serial sdAb dilutions in PBS starting from 500 nM were added. All incubations were carried out for 1 h at RT and after every incubation, plates were washed four times with PBS. The fluorescently labelled sdAbs were detected directly, or via indirect detection using an incubation with rabbit anti-VHH (clone QE19, 1:2000; QVQ BV, Utrecht, The Netherlands), followed by an IRDye800conjugated goat-anti-rabbit antibody (1:2000, LI-COR Biosciences, Lincoln, NE, USA). The plate was scanned using the Odyssey near-infrared scanner (LI-COR Biosciences, Lincoln, NE, USA).
2.7.2. On Cells
SCCVII or SCCVII-rPDGFRβ cells (approximately 104 cells per well) were seeded in a Nunclon™ Delta Surface 96-wells tissue culture plate (Thermo Fisher Scientific, Bleiswijk, The Netherlands) and allowed to adhere overnight. Three-fold serial sdAb dilutions were added onto cells in binding medium (DMEM without phenol red, supplemented with 25 mM HEPES, 1% w/v bovine serum albumin, pH 7.2) at 4 °C to prevent internalization. After 2 h of incubation, the sdAb was removed and cells were fixed by incubating the cells in 4% PFA for 20 min at RT. Detection of sdAbs was performed as described above.
2.8. Competition Assay on Immobilized Ectodomain
Purified rat PDGFRβ (Sino-Biological, Eschborn, Germany) was coated overnight at 4 °C at 1 μg/mL in PBS in ELISA plates (Nunc MaxiSorp™, Thermo Fisher Scientific, Bleiswijk, The Netherlands). Next day, plates were washed with PBS and blocked with PBS, supplemented with 4% (w/v) skimmed milk powder (Merck Millipore, Darmstadt, Germany) and 0.05% v/v Tween, for 1 h at RT. All further incubations were carried out for 1 h at RT in PBS supplemented with v/v 0.05% Tween (PBST). A fixed concentration of IRDye800 conjugated sdAb was mixed with unconjugated competitors (sdAbs or ligand) in three-fold serial dilutions, starting with a 20-fold molar excess. Fluorescent signal of the IRDye800 conjugated sdAb was detected using the Odyssey near-infrared scanner (LI-COR Biosciences, Lincoln, NE, USA).
2.9. Internalization Assay
SCCVII-rPDGFRβ cells (approximately 10
4 cells per well) were seeded in a 96-wells tissue culture plate (Nunclon™ Delta Surface, Thermo Fisher Scientific, Bleiswijk, The Netherlands) and allowed to adhere overnight. Next day, internalization of the IRDye800 conjugated sdAb (5 nM) into the cells was measured after an incubation at 37 °C for period of 15 min. Total fluorescence as well as fluorescence attributed to the internalized fraction were measured and used to calculate the internalization rate constant as described by Heukers et al. [
22]. The internalized fraction is assessed by performing two subsequent acid washes (0.2 M glycine-HCl, 150 mM NaCl, pH 2.3) removing the membrane bound sdAb fraction from the cells. For the cell loading assays, the same procedure was applied with minor modifications. Longer timepoints (up to 72 h) were measured and only the internalized fraction was plotted.
2.10. PDGFRβ Activation Assay
SCCVII-rPDGFRβ cells (approximately 105 cells per well) were seeded in a 12-wells tissue culture plate (Nunclon™ Delta Surface, Thermo Fisher Scientific, Bleiswijk, The Netherlands) and allowed to adhere overnight. The next day, the medium was refreshed with medium containing 10 ng/mL of the PDGFRβ ligand (PDGF-BB) and/or 10 nM PDGFRβ specific sdAbs. After 15 min of incubation at 37 °C, cells were cooled down on ice and washed twice with ice-cold PBS. Cell lysates were prepared by taking the cells up in 30 μL 1× Laemmli protein sample buffer without DTT. Next, 20 µL lysates were boiled for 10 min at 100 °C, loaded on a 8–12% (w/v) PAGE gel (Bio-rad, Veenendaal, the Netherlands) and blotted onto a PVDF membrane (Roche, Mannheim, Germany). Blots were stained overnight for phosphorylated PDGFRβ using a rabbit polyclonal antibody targeting the Tyr751 phosphorylated receptor (Cell Signaling Technology, Leiden, The Netherlands), followed by a 1 h incubation with IRDye800-conjugated goat-anti-rabbit secondary antibody (LI-COR Biosciences, Lincoln, NE, USA). As a loading control, the blots were also stained for actin using a monoclonal mouse-anti-actin antibody (Sigma-Aldrich, Zwijndrecht, The Netherlands), followed by a 1 h incubation with an IRDye680-conjugated goat-anti-mouse secondary antibody (LI-COR Biosciences, Lincoln, NE, USA). Fluorescent signal was detected using the Odyssey near-infrared scanner (LI-COR Biosciences, Lincoln, NE, USA).
2.11. Confocal Fluorescence Microscopy
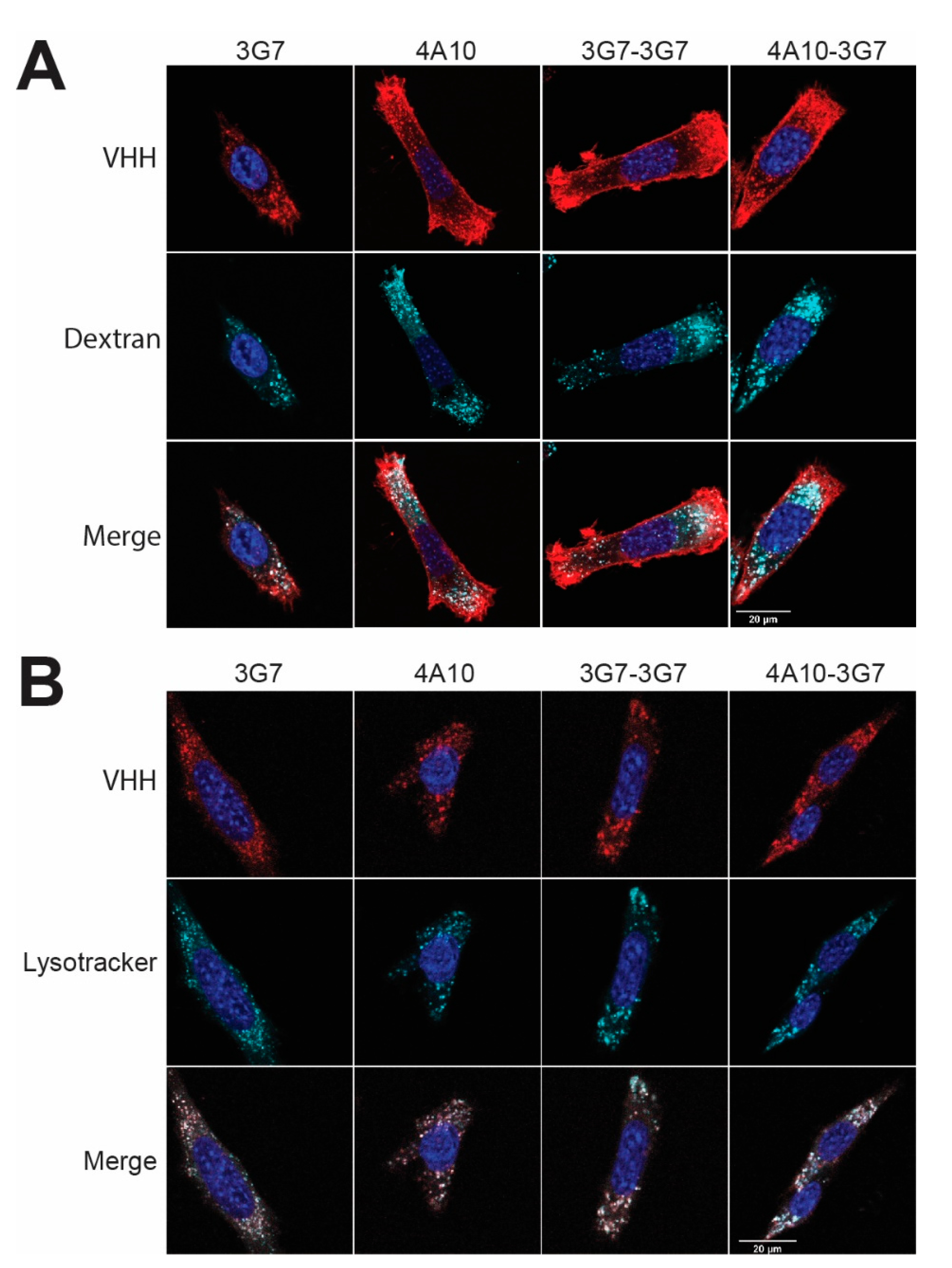
SCCVII-rPDGFRβ cells were seeded on cover slips in a 24-wells tissue culture plate (approximately 104 cells per well) and allowed to adhere overnight. The next day, cells were incubated for 2 h at 37 °C with 5 nM AlexaFluor647-conjugated sdAbs in binding medium (DMEM without phenol red, supplemented with 25 mM HEPES, 1% w/v bovine serum albumin, pH 7.2). 60 nM lysoTracker™ Red DND-99 (ex 577 nm, em 590 nm) (Thermo Fisher Scientific, Bleiswijk, the Netherlands) or 1 mg/mL TMR-Amino Dextran 70 kDa (ex 555 nm, em 580 nm) (Fina Biosolutions LLC, Rockville, MD, USA) were co-incubated with the AlexaFluor647-conjugated sdAb conjugates (ex 650 nm, em 668 nm). Upon incubation, cells were washed with binding medium and PBS. Afterwards, the cells were fixed in 4% PFA and PFA-induced autofluorescence was quenched with 50 mM glycine in PBS for 15 min at RT. Nuclei were stained with DAPI (ex 358 nm, em 461 nm) (Thermo Fisher Scientific, Bleiswijk, the Netherlands) for 5 min and after washing, the cover slips were mounted using Mowiol (Merck Millipore, Darmstadt, Germany). Images were taken using a Zeiss LSM700 confocal microscope (Carl Zeiss Microscopy GmbH, Germany) equipped with a 63× oil immersion objective. Three images were taken, after which a representative was included in the figure. All images are taken with the same microscope settings and no image editing has been performed.
2.12. Mass Spectometry Analysis of sdAb-Auristatin F Conjugates
LC-MS analysis was performed using a Thermo Finnigan LC system (Thermo Finnigan, San Jose, CA, USA) coupled to a Bruker Q-TOF mass spectrometer (Bremen, Germany) equipped with an electrospray ionization (ESI) source. Mass determination was performed using a Zenix-C column (4.6 × 300 mm; 5 μm; Sepax Technologies Inc., Newark, DE, USA). The mobile phase consisted of a mixture of water, acetonitrile, trifluoroacetic acid and formic acid (79.9/19.9/0.1/0.1, v/v/v/v, respectively). A 17-min isocratic run was performed, and MS analysis was achieved in positive ionization mode. The protein ion charge assignment and molecular mass determinations were performed using the “Charge Deconvolution” utility of Bruker Daltonics Data Analysis software.
2.13. Cell Viability Assays
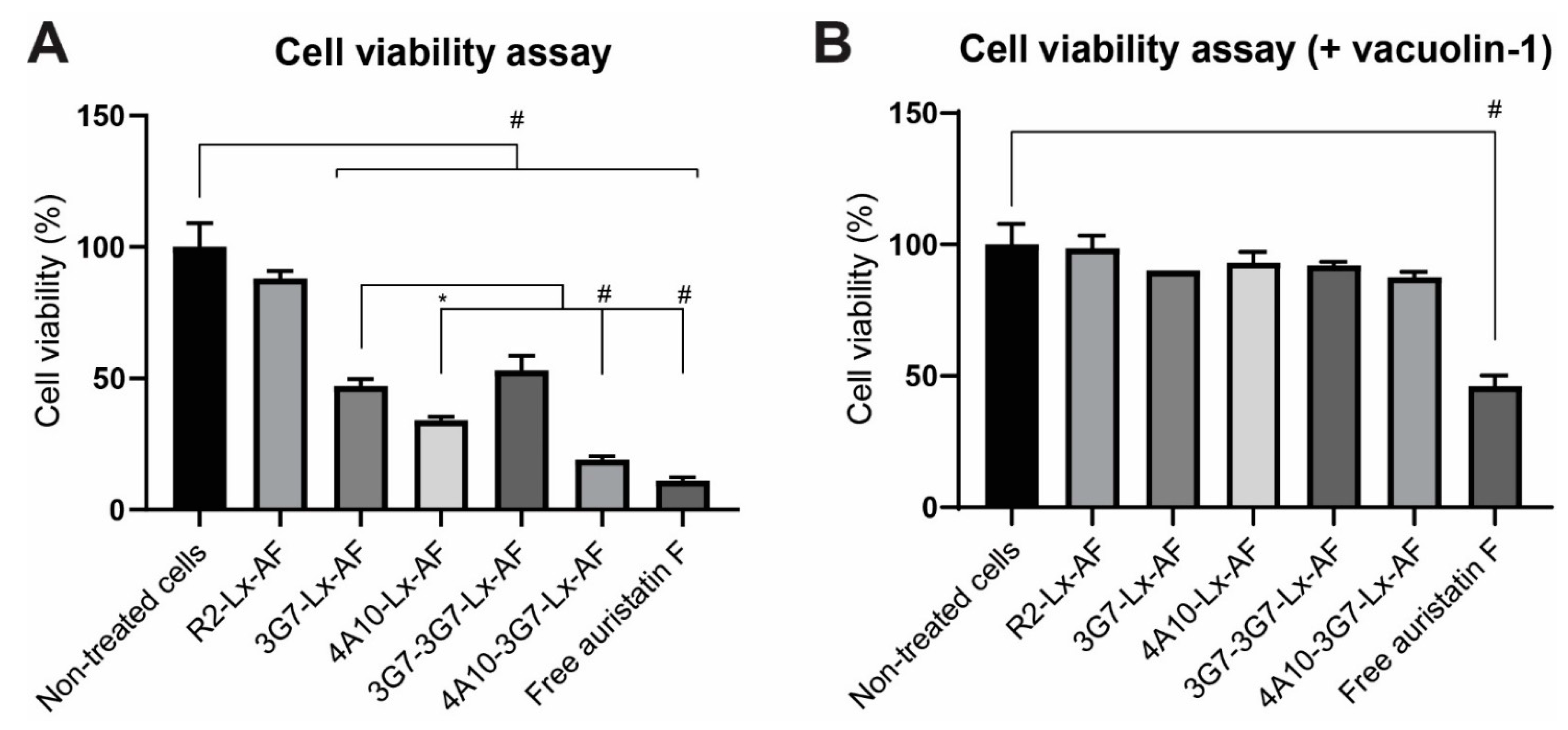
Cell viability assays were performed using the RealTime-Glo™ MT Cell Viability assay (Promega, Leiden, The Netherlands) according to manufacturer’s protocol or using the CellTiter-Blue® assay (Promega, Leiden, The Netherlands) according to manufacturer’s protocol. As starting points, 750 cells/well were seeded in an opaque walled white 96-well plate to maintain assay linearity. SCCVII or SCCVII-rPDGFRβ were incubated with different sdAb-Lx-Auristatin F constructs and monitored for 72 h by measuring the luminescence or fluorescence using the FLUOstar OPTIMA FL microplate reader (BMG LABTECH). When the cell viability assay was performed in the presence of lysosomal exocytosis inhibitor vacuolin-1, the same procedure was applied with minor modifications. In a separate experiment cells were pre-incubated for 3 h with 1 µM vacuolin-1 (Sigma-Aldrich, Zwijndrecht, The Netherlands), after which the medium was refreshed with medium containing the sdAb-drug conjugates and 1 µM vacuolin-1.
2.14. Receptor Quantification on SCCVII-rPDGFRβ Cells
A binding assay on cells and a titration series of IRDye800 conjugated PDGFRβ binding sdAb (3G7) was performed in parallel. The binding assay on cells was performed as described before, with the modification that the cells were lysed in 100 µL RIPA buffer before the plate was scanned.
In the titration series, a concentration range of the IRDye800 conjugated sdAb was made starting at 10 nM and ending at 1.2 × 10−3 pM in a volume 100 µL. The Bmax of the binding assay was interpolated with the obtained titration series to calculate the number of fluorescently bound molecules per well. To determine the average number of cells per well, cells were harvested and counted. The number of fluorescent molecules per well was then divided by the number of cells per well to obtain the number of fluorescent molecules per cell and thus the number of receptors localized on the cell membrane per cell.
2.15. Live Cell Imaging Using Spinning Disc Microscopy
SCCVII-rPDGFRβ cells were seeded in a glass bottom µ-slide 8-well chambered coverslip (Ibidi, Gräfelfing, Germany) (approximately 104 cells per well) and allowed to adhere overnight at 37 °C, 5% CO2 in a humidified incubator. The next day, the cells were transferred the TokaiHit incubation chamber of the microscope to keep the cells at 37 °C. At the starting point of the timelapse-experiment, 10 nM sdAb conjugated to maleimide pHrodo™ Red (Thermo Fisher Scientific, Bleiswijk, The Netherlands) was added to the cells. Live-cell imaging was performed using a using a Nikon Eclipse Ti confocal spinning disc microscope equipped with Perfect Focus System and a 60× oil objective. Metamorf software was used to make a time-lapse video (picture every 15 sec with 250 ms exposure time). Acquired files were background corrected and time stamped with Fiji/ImageJ software.
4. Discussion
Recent studies are encouraging the development of smaller antibody-drug conjugates or sdAb-drug conjugates for their advantage in tissue distribution, and especially homogeneous targeting [
12,
13]. In this proof-of-principle study we investigate a panel of sdAbs targeting the same receptor but with different internalization rates to determine whether this correlates with the overall efficacy of intracellular drug delivery in vitro.
Previous studies have shown that the internalization rate of antibodies can be affinity dependent [
25,
26]. In vitro internalization assays revealed that antibody internalization generally increased with affinity, plateauing once the rate of antigen internalization exceeded the rate of antibody dissociation [
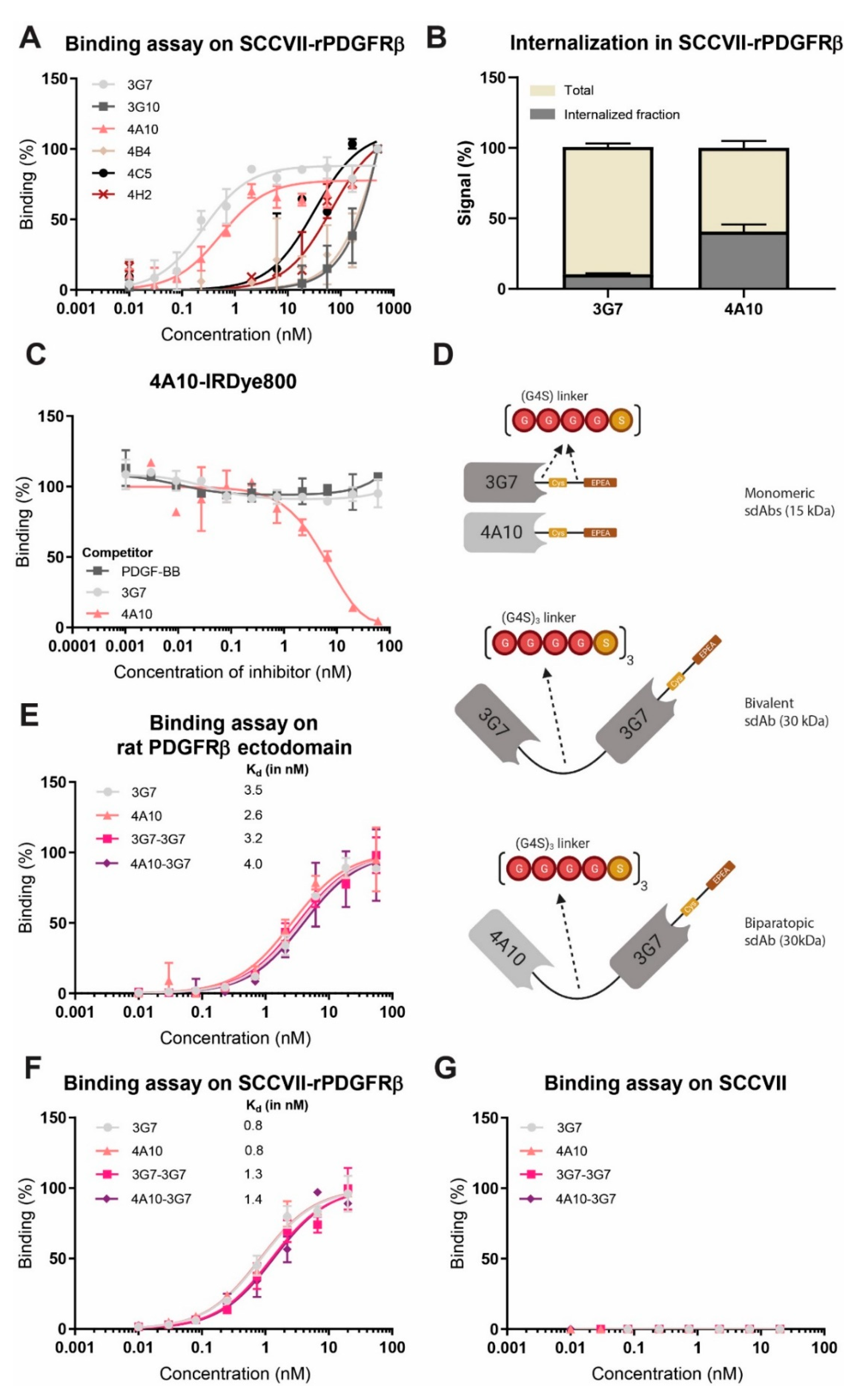
25]. In our study, as all investigated constructs have similar binding affinities, in the low nanomolar range, it is not expected that this will affect internalization (
Figure 2E,F).
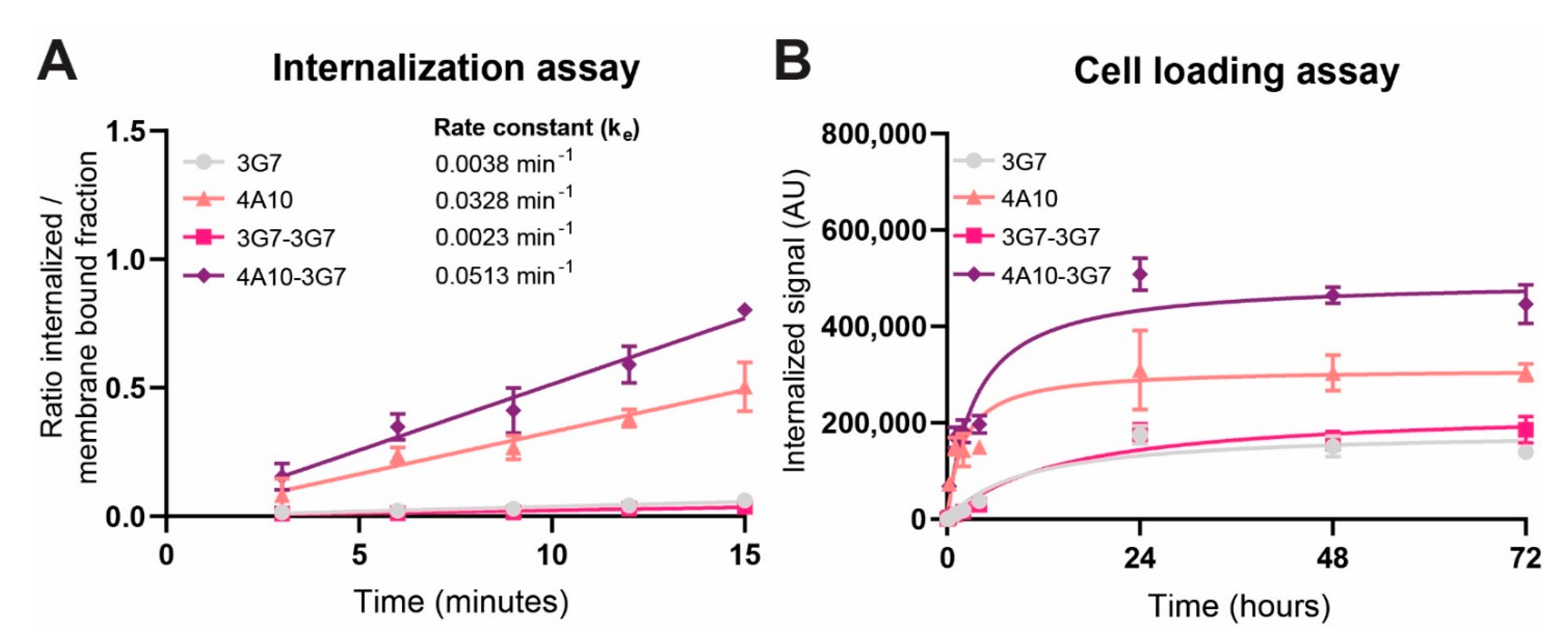
We tested internalizing monomeric, bivalent and biparatopic sdAb constructs and showed that biparatopic construct 4A10-3G7 clearly internalizes fastest, followed by the monomer 4A10. The internalization rate of these fast internalizing constructs even seem to be slightly higher than the reported internalization rate of the natural PDGFRβ ligand PDGF-BB, known to dimerize the receptor [
27]. Monomer 3G7 and bivalent construct 3G7-3G7 internalize relatively slow compared to the other sdAb constructs. The difference in internalization speed between both monomers could indicate that internalization speed is a characteristic of the targeting moiety, possibly depending on the binding epitope, and not solely on the target receptor.
An interesting observation is that addition of a slow internalizing sdAb to a fast-internalizing sdAb led to an even faster internalizing sdAb construct, biparatopic 4A10-3G7. In a recent study, a fast internalizing single-chain variable fragment (scFv) fused to a non-binding antibody was rendered slow internalizing by substitution of the non-binding antibody for a slow-internalizing antibody [
28]. Based on this example, one could expect that the biparatopic sdAb construct would internalize slower than 4A10 alone. However, here the two sdAbs combined lead to increased internalization rates.
Other studies have indicated that bivalent sdAbs may induce receptor dimerization, while biparatopic sdAbs may induce receptor dimerization and/or oligomerization, leading to faster receptor internalization [
22,
29]. 4A10-3G7 could induce PDGFβ receptor oligomerization leading to faster internalization speed than 4A10 alone. It may be that 3G7-3G7 cannot induce receptor dimerization due to the linker length of the bivalent construct or the binding epitope of 3G7. This could explain the similar internalization speed as the monomeric construct 3G7 and substantiates our suggestion that the binding epitope of the targeting moiety can influence the internalization rate.
Importantly, we show that the internalization rate reflects the intracellular accumulation after a longer period of time: targeting moieties with higher internalization rates transport more cargo into the cell. That excludes the efficient recycling back to the membrane, which has for instance been described for HER2-targeted ADC Kadcyla [
30]. This is in line with other studies, including one study in which surface depletion of the fast internalizing target antigen is observed [
28]. This may also be an explanation for the stabilization of the intracellular accumulation we observed.
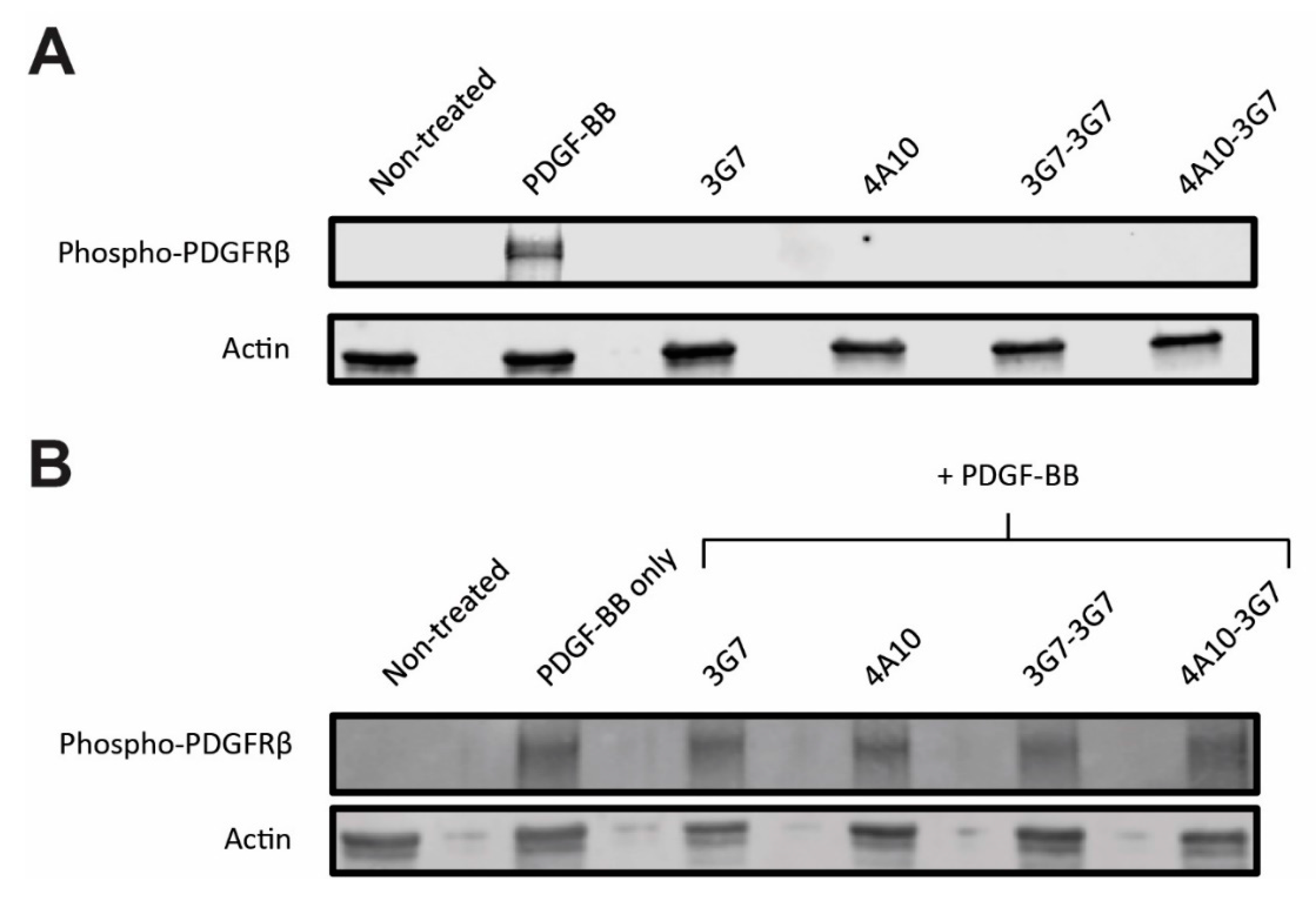
When the internalization kinetics of the different sdAb constructs was investigated, it was determined that their binding does not interfere with the activation status of the PDGFβ receptor. We identified a possible route of sdAb internalization, showing that all sdAbs internalize mainly via macropinocytosis and end up in the lysosomes, the ideal destination for intracellular drug delivery [
3,
4,
31]. Macropinocytosis of the PDGFRβ complex has been described before, but here macropinocytosis is induced without phosphorylating the receptor [
32,
33].
Conjugated to the cytotoxic payload Auristatin F (AF), the sdAb-Lx-AF-conjugates with faster internalization rates displayed more cytotoxicity than slower internalizing AF-conjugates. In other studies fast internalizing sdAb constructs were also more potent in vitro, although not much attention has been given to these findings before [
12,
13]. When the same assay was performed in presence of vacuolin-1, a lysosomal exocytosis inhibitor [
24], none of the conjugates showed any cell killing. This shows that the sdAbs deliver drugs via the endosomal-lysosomal degradation pathway, making them interesting candidates for intracellular drug delivery [
34].


 and
and 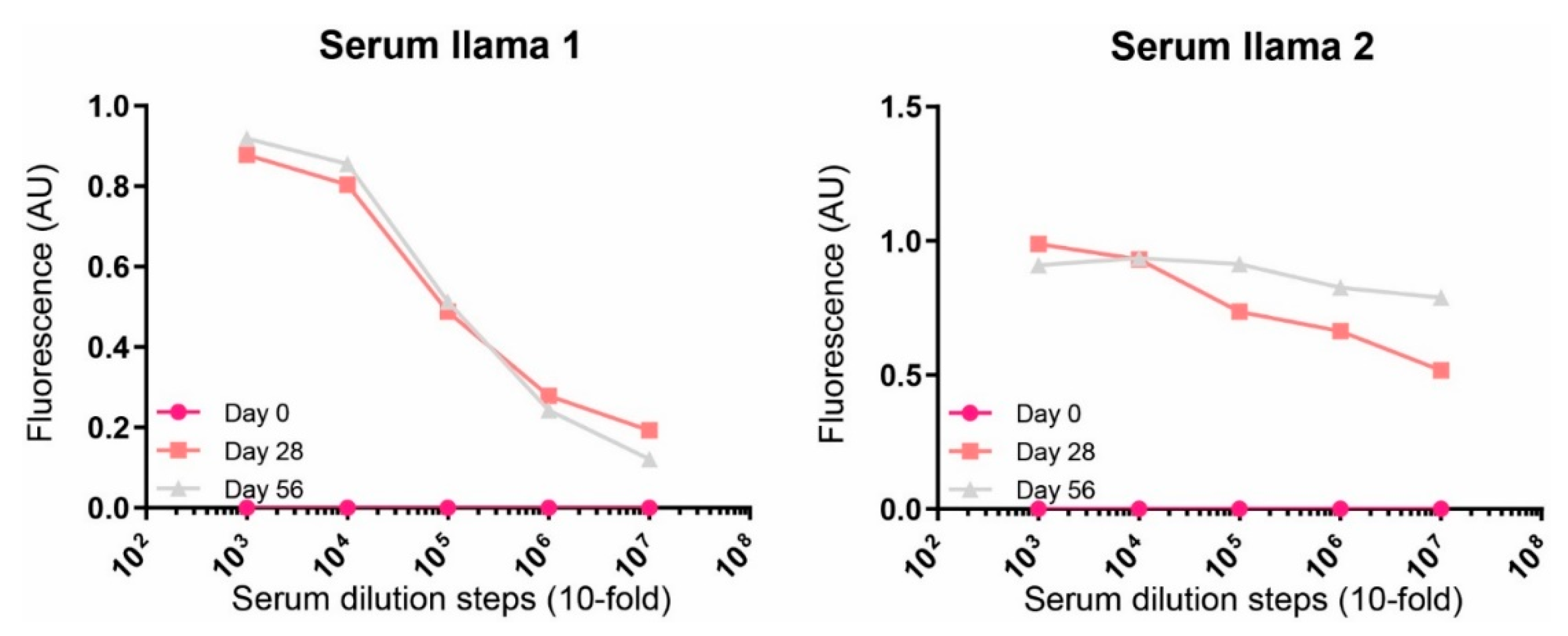{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





